Archeocéty: starověké velryby a počátky kytovců (Eocén–Oligocén)
Archeocéty — příběh starověkých velryb (Eocén–Oligocén): od obojživelných forem k předkům delfínů a plejtváků. Evoluce, fosilie a migrace z Indopákistánu do světa.
Archaeoceti („starověké velryby“), ve starší literatuře nazývané Zeuglodontes, představují skupinu raných a primitivních kytovců. Existovaly od raného eocénu až do pozdního oligocénu (přibližně před 55–23 miliony let). Byly první rozsáhlou evolucionární radiací kytovců a zahrnují mnohá přechodná, částečně obojživelná stádia ve vývoji kytovců. Archeocéty tedy stojí u kořenů obou moderních podřádů, mysticeti (velryby s filtrovacími destičkami) a odontoceti (zubozobí kytovci).
Galerie obrázků
10 Obrázky







Časové rozmezí a rozšíření
Archeocéty vznikly v mělkých pobřežních vodách oddělujících Indii a Asii v období asi 55–45 mya. V raném eocénu (přibližně 55–48 mya) pocházejí známé fosilie téměř výhradně z indopákistánské oblasti. Do středního až pozdního eocénu (41–34 mya) se jejich fosilie objevují téměř po celém světě — včetně Severní Ameriky, Egypta, Nového Zélandu a Evropy.
Protože archeocéty nebyly tak dokonale přizpůsobené životu na otevřeném oceánu jako dnešní kytovci, šířily se především podél pobřeží. Do Severní Ameriky se pravděpodobně dostaly pobřežními trasami — kolem Eurasie přes moře Tethys a podél pobřeží Evropy, Grónska až k americkým břehům — spíše než přímým přechodem otevřeným oceánem.
Anatomie a adaptace
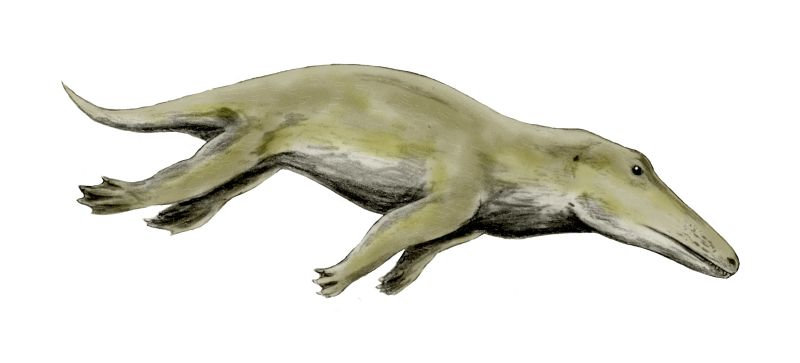
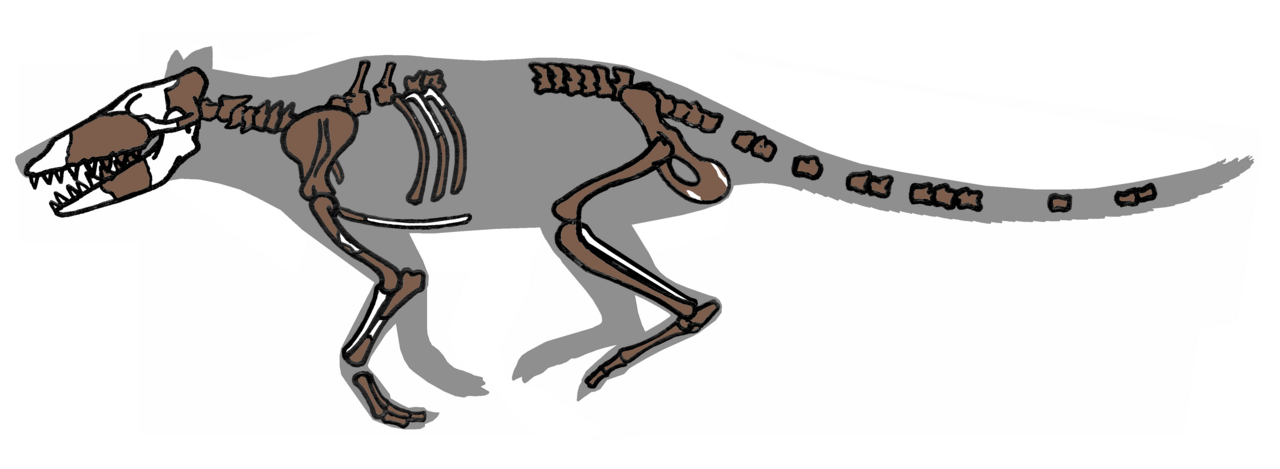
Archeocéty vykazují směs terestrických a adaptací na vodní život. Mezi důležité anatomické znaky patří:
- Lebka a sluch: přítomnost charakteristického zesílení v oblasti středního ucha (involucrum), které je diagnostické pro kytovce a napomáhalo vedení zvuku ve vodě;
- Zuby: plně vybavený zubní aparát (heterodontní zuby u starších forem), což je rozdíl oproti filtrujícím místům u pozdějších mysticetů;
- Nůžky a nosní otvor: postupná migrace nosních otvorů směrem dozadu na lebce, což je předstupeň vzniku oficiálního „dechu“ (blowhole) u moderních kytovců;
- Končetiny a pohyb: rané rody (např. Pakicetus) měly dobře vyvinuté končetiny a byly schopné pohybu po souši, středně pokročilé formy (např. Ambulocetus) plavaly pomocí pádlovitých končetin a vertikálního vlnění těla; pokročilé mořské formy (např. Basilosaurus, Dorudon) měly zmenšené zadní končetiny a pohybovaly se převážně pomocí svislého vlnění trupu a ocasu, někdy již s patrnou ocasní ploutví;
- Velikost: rozmanitost velikostí od menších forem několik desítek kilogramů až po velmi velké Basilosaurus (délka přes 15–18 m).
Ekologie a způsob života
Archeocéty obývaly převážně mělká pobřežní a příbřežní prostředí, estuária a laguny, kde lovily ryby a jiné mořské obratlovce. Některé druhy byly částečně obojživelné a využívaly i souš pro odpočinek či rozmnožování. Jejich zuby a čelistní stavba naznačují aktivní predátorskou strategii — uchopení a drcení kořisti — na rozdíl od pozdějších filtrujících nebo echolokačních strategií.
Hlavní rody a významné fosilní nálezy
- Pakicetus — raný, převážně suchozemský rod, ukázal, že předci kytovců žili na souši;
- Ambulocetus — „chodící velryba“, dobře přizpůsobená jak plavání, tak chůzi po břehu;
- Rodhocetus, Protocetus, Georgiacetus — středně pokročilé formy ukazující postupné ztrácení závislosti na souši a zlepšování plaveckého způsobu pohybu;
- Basilosaurus — plně mořský, hadovitě prodloužený trup, velké rozměry a malé, zakrnělé zadní končetiny;
- Dorudon — relativně malý, plně mořský archeocét souběžný s Basilosaurusem, častý v eocénních sedimentech; často uváděn jako blízký příbuzný předků moderních kytovců.
Vývojové souvislosti a filogeneze
Skupina je považována za parafyletickou, protože z ní vycházejí oba moderní podřády kytovců. Archeocéty představují první velkou evoluční vlnu kytovců; moderní znaky, jako je specializovaná echolokace u odontocetů nebo filtrační potrava u mysticetů, se však vyvinuly až později, během druhé radiace kytovců přibližně před 36–35 mya.
Závěr
Archeocéty dokumentují klíčový evoluční přechod od suchozemských savců k plně mořským kytovcům. Díky bohatému fosilnímu záznamu — od těžkopádných suchozemských forem přes polopobřežní „chodící velryby“ až po plně mořské Basilosauridae — poskytují detailní obraz anatomických a ekologických změn, které vedly ke vzniku dnešních velryb a delfínů.
Otázky a odpovědi
Otázka: Co jsou to archeoceti?
Odpověď: Archaeoceti, ve starší literatuře známí také jako Zeuglodontes, je skupina primitivních kytovců, kteří žili od raného eocénu do pozdního oligocénu, tedy před 55-23 miliony let.
Otázka: Jaké dva moderní podřády pocházejí z Archaeoceti?
Odpověď: Dva moderní podřády, které vzešly z Archaeoceti, jsou Mysticeti a Odontoceti.
Otázka: Kde se Archaeoceti vyvinuli?
Odpověď: Archaeoceti se vyvinuli v mělkých vodách, které před 55-45 miliony let oddělovaly Indii a Asii.
Otázka: Kolik druhů archeocetů bylo nalezeno?
Odpověď: Bylo nalezeno asi 30 druhů archeocetů, kteří byli přizpůsobeni plně oceánskému životu.
Otázka: Kdy se u kytovců vyvinula echolokace a filtrování potravy?
Odpověď: Echolokace a filtrační krmení se vyvinuly později, během druhé radiace před 36 až 35 miliony let.
Otázka: Kde se nacházely rody archeocetů před 41-34 mil. let?
Odpověď: V období 41-34 mya byly rody archeocetů známy po celé Zemi, včetně Severní Ameriky, Egypta, Nového Zélandu a Evropy.
Otázka: Jak se dostali do Severní Ameriky?
Odpověď: Je velmi nepravděpodobné, že by byli tak dobře přizpůsobeni cestování po otevřeném oceánu jako žijící kytovci, takže se předpokládá, že se do Severní Ameriky dostali po pobřežních vodách buď kolem Afriky, nebo přes moře Tethys (mezi Eurasií a Afrikou) a podél pobřeží Evropy, Grónska a Severní Ameriky.
Štítky
Související články
Autor
AlegsaOnline.com Archeocéty: starověké velryby a počátky kytovců (Eocén–Oligocén) Leandro Alegsa
URL: https://cs.alegsaonline.com/art/5276
Zdroje
- paleodb.org : "Archaeoceti"
- annualreviews.org : annualreviews.org/doi/abs/10.1146/annurev.ecolsys.33.020602.095426
- digitallibrary.amnh.org : digitallibrary.amnh.org/dspace/handle/2246/5671